| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
| ||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||
Correspondence to: Aart J. E. van Bel, aart.v.bel@bot1.bio.uni-giessen.de (E-mail), 49-641-99-35119 (fax).
|
|
ABSTRACT |
|---|
A method was designed for in vivo observation of sieve element/companion complexes by using confocal laser scanning microscopy. A leaf attached to an intact fava bean plant was mounted upside down on the stage of a confocal microscope. Two shallow paradermal cortical cuts were made in the major vein. The basal cortical window allowed us to observe the phloem intact. The apical window at 3 cm from the site of observation was used to apply phloem-mobile fluorochromes, which identified living sieve elements at the observation site. In intact sieve tubes, the sieve plates did not present a barrier to mass flow, because the translocation of fluorochromes appeared to be unhindered. Two major occlusion mechanisms were distinguished. In response to intense laser light, the parietal proteins detached from the plasma membrane and formed a network of minute strands and clustered material that aggregated and pressed against the sieve plate. In response to mechanical damage, the evenly distributed P plastids exploded, giving rise to the formation of a massive plug against the sieve plate. In case of mechanical damage, the parietal proteins transformed into elastic threads (strands) that extended throughout the sieve element lumen. Our observations cover the phenomena encountered in previous microscopic and electron microscopic studies and provide a temporal disentanglement of the events giving rise to the confusing mass of structures observed thus far.
|
|
INTRODUCTION |
|---|
Since the discovery of sieve elements (SEs) by Theodor ![]()
![]()
![]()
![]()
One of the stumbling blocks to full acceptance of pressure flow as the mechanism of phloem translocation is the typical components of the SEs. In particular, those directly behind the sieve plates may obstruct mass flow. These bodies, originally designated as Schleim (slime) by ![]()
![]()
![]()
![]()
![]()
In electron microscopic images, a parietal filamentous network that fully or partly plugged the sieve pores was observed (![]()
![]()
![]()
![]()
![]()
![]()
The debate between the proponents of mass flow and those who claimed the existence of structural barriers to mass movement prompted massive research efforts (for an overview, see ![]()
![]()
![]()
![]()
![]()
![]()
![]()
The ongoing discussion on the mechanism of phloem transport essentially hinges on one question. What is the in vivo structure of SEs? With reference to the occlusion of the sieve pores, many investigators have pointed out that the main obstacle to elucidate the in vivo structure of SEs is their extreme sensitivity to any kind of manipulation (e.g., wounding, cutting, or chemical fixation). The uncertainties with regard to SE ultrastructure have left their mark in the handbooks. The drawings are basically very indefinite, with the exception of the one in ![]()
Our investigations have dealt with the visualization of the structure of undamaged and actually translocating SEs by using confocal laser scanning microscopy (CLSM). Use of various fluorescent dyes enabled us to observe intact SEs in whole plants mounted on the stage of the confocal laser scanning microscope. This study shows that intact SEs are essentially devoid of occlusions in the sieve pores and presents new insights into the origin, time scale, order of emergence, and triggering of the occlusion mechanisms.
|
|
RESULTS |
|---|
Fluorochromes Brought into Action
A variety of fluorochromes was employed for confocal imaging of the structural components of the SEs in tissue slices and in actually transporting (i.e., living) sieve tubes in intact plants. For identification of intact SEs, membrane-permeant, phloem-mobile fluorescent probes were used. The general advantage of membrane-permeant fluorochromes is to avoid microinjection techniques that may inflict major cell damage ( ![]()
Two of the phloem-mobile fluorochromes brought into action belong to the 5(6)carboxyfluorescein diacetate (CFDA) family. These dyes have in common that they cross the plasma membrane in acetate form and are cleaved by cytosolic enzymes producing membrane-impermeant carboxyfluoresceins (handbook from the manufacturer; Molecular Probes, Eugene, OR). CFDA has been used extensively in plant research as a marker for symplasmic transport (![]()
![]()
![]()
![]()
![]()
The nonpolar RH dyes have been used for membrane staining in neurophysiological studies (e.g., ![]()
|
CLSM of SEs in Tissue Slices; Identification of SE Components
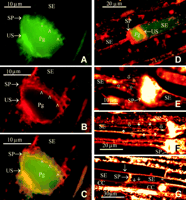
In a first series of experiments, longitudinally cut slices of phloem tissue were treated with CDCFDA/RH-160 mixtures. The confocal laser scanning microscopic images of the slices were compared with those obtained with conventional light microscopy and transmission electron microscopy. The comparison aided in the identification of the cellular structures emerging in confocal images.
In keeping with the behavior of CFDA (![]()
![]()
![]()
![]()
![]()
The nonpolar RH-160 turned out to be an excellent plasma membrane marker. The fluorochrome instantly associated with the plasma membrane of intact cells and to most membranous structures within the cells after 30 to 120 min (Figure 1E to G). Among them are the tonoplast and the membranous structures of chloroplasts, mitochondria (Figure 1G), and P plastids. Apart from the "established" membranous structures, vesicles of undetermined composition expanding in the SEs in response to injury (Figure 1F) were stained by RH-160.
As reported earlier (e.g., ![]()
![]()
In SEs, somewhat deeper inside a tissue slice, some strands extended through the cytosolic lumen from the one sieve plate to the other, whereas most strands branched away to the side walls (Figure 1F). The first species of strands may be the ones reported by ![]()
![]()
Visual observation of the middle part of the tissue by CLSM (up to 50 to 100 µm, depending on the overlaying cell types) showed the absence of strands or vesicles in SE/CC complexes that were away from the transverse cuts. In these apparently less damaged SEs, a few P plastids pressed against the sieve plate were present, and the SE plasma membrane was locally detached from the cell wall (Figure 1G).
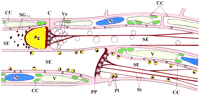
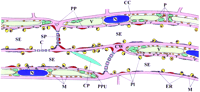
Nature and position of several SE components may be artifactual (i.e., subject to wounding responses). This probably holds true for most of the electron microscopic images published (artifact-inducing cutting precedes fixation). A tentative reconstruction of the wound reaction in successive SEs was made. This reconstruction was based on a broad range of observations in CDCFDA/RH-160-stained tissue slices. The diagram in Figure 2 (top) shows the structure of the SEs imposed by different degrees of injury. Triggering of the diverse structural reactions is discussed when the insights gained from the injury experiments are taken into consideration.
|
How to Observe Intact SEs
As outlined in the Introduction, the in vivo structure is foremost for the assessment of the physiological potential of the sieve tubes. Because a paramount interest lies in the functioning of the phloem in intact plants, we designed a method to observe the SEs intact.
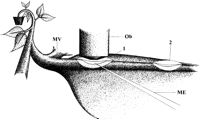
Cell sheets that were a few layers thick were sliced paradermally at two locations 3 cm apart from each other. Cell sheets were removed from the lower side of the main vein of mature broad bean leaves still attached to an intact plant (Figure 3). The two-site removal of overlaying tissues without disturbing the phloem allowed us to apply phloem-mobile fluorochromes in the vicinity of the SEs at the one site and to observe tracer translocation through living SEs by CLSM at the other (Figure 3). Use of the special high-magnification water immersion objective for CLSM had essential advantages. It enabled close visual access to the SEs without a glass cover that is necessary for oil immersion. The cover slip would also damage the surrounding tissues. In addition, the working distance of 220 µm of the water immersion objective permitted the manipulation and impalement of microelectrodes under high-resolution surveillance.
|
At the site of application (Figure 3, position 2), the phloem-mobile tracers accumulated in all cell types but mainly in the SE/CC complexes. However, at the site of observation (Figure 3, position 1), fluorescence was only visible within the SE/CC complexes. The two-site approach made the SE/CC complexes stand out against the other cells and identified transporting (i.e., living) SEs. The direction of the dye movement was checked; CFDA only moved basipetally in the phloem (data not shown). In conclusion, CFDA was translocated in SEs, which are fully functional and most likely displaying the in vivo structure.
CFDA translocation usually needed 20 to 30 min to cover the distance of ~3 cm between the site of application and the site of observation. In this time interval, the fluorochrome must first have diffused through the apoplast of the overlying cells to the SE plasma membrane. After having crossed the plasma membrane of the SEs in the acetate form, CFDA was presumably hydrolyzed by esterases inside of the SEs (![]()
![]()
|
In Vivo Structure of the SE/CC Complex
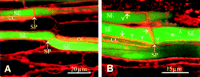
Optical sectioning at two levels through the same vascular bundle underlined that CFDA is moving exclusively in the sieve tubes (Figure 4C and Figure 4D). Translocated CFDA obviously remained restricted to the SE/CC complexes (Figure 4A to D); no fluorescence unloading into the adjoining cell elements was detected. Apart from the starch grains in the P plastids, visible as minute bright spots in the SEs (Figure 4C and Figure 4D), none of the cellular components of the SE/CC complexes was stained by CFDA. Detailed pictures of actually transporting SEs showed a homogeneous distribution of CFDA in the SEs and a stronger but unevenly distributed accumulation of fluorochrome inside of the CCs (Figure 4C and Figure 4D). The dye preferentially accumulated in the lobed vacuole of the CCs, demarcating the location of the nuclei. The images illustrate the coincident ending of SEs and CCs and the staggered position of the CCs (Figure 4C and Figure 4D), as was observed for SE/CC complexes of Lupinus and Vicia labeled with intracellularly injected LYCH (![]()
![]()
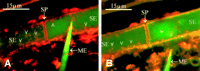
After 15 min of staining the phloem with RH-160 at the site of observation, the membrane lining of the SEs was clearly visible (Figure 4E). The SEs appeared to be devoid of the subcellular structures (Figure 4E), as observed in injured tissues (Figure 1E and Figure 1F); plugs, strands, and vesicles were absent. The plasma membrane of the SE could be observed crossing the sieve pores (Figure 4E). The pore plasmodesma units (Figure 4E) were more fluorescent at the CC side (the branched side with more plasma membrane material) than at the SE side (the single-pore side).
After prolonged treatment (1 hr) with RH-160, stained particles emerged inside the SEs, which implies that RH-160 is membrane permeant (Figure 4F). In the CCs, numerous small (probably mitochondria) and larger (probably chloroplasts) particles were visible (Figure 4E and Figure 4F). In the SEs, parietal bodies (probably P plastids) showed up which are evenly distributed pressed against the plasma membrane (Figure 4F, Figure 5B, and Figure 6A).
|
|
Besides the P plastids, unequally distributed plaques of fluorescent sediment are parietally located (Figure 4F, Figure 5A, Figure 5B, and Figure 6A). It appeared that RH-160 (Figure 4F and Figure 5B) and CDCFDA (Figure 5A) labeled the same sediment. The sieve plate is largely free from sediment, apart from scattered aggegrations, as shown by double-staining experiments in which the SEs were distantly labeled (by applying at position 2 in Figure 3) with HPTSA and locally labeled (position 1 in Figure 3) with RH-160 (Figure 5B). The massive CDCFDA-tagged plugs, which were observed on sieve plates in tissue slices (Figure 1D), were absent in the intact SEs (Figure 4E, Figure 4F, Figure 5A, and Figure 5B). Occasionally, a thicker fluorescent sediment on the sieve plate appeared (Figure 4F and Figure 5B). This sedimentary layer did not seem to prevent the passage of the phloem-mobile HPTSA (Figure 5B). Use of HPTSA revealed the existence of large spindle-shaped clusters of material (probably nondispersed crystalline P protein bodies) inside of the living SEs (Figure 5B).
Effects of Laser Light on SE Structure
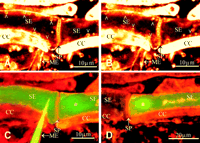
Because the mass of sediment on the sieve plates in intact SEs seemingly correlated with the length of the observation period, the effect of laser light on the deposition of material was studied. A CLSM time scan of an intact SE stained with RH-160 showed dramatic structural responses to an increase of the laser intensity (Figure 6A to D). After raising the laser energy flow rate from ~45 to 100 µW, parietal packets of stained material detached from the cell wall within a few seconds and produced opaque structures in the cytosolic lumen (Figure 6A and Figure 6B). The particles all moved in the same direction (Figure 6C), apparently being dragged along with the mass flow inside of the SE. Within 1 min, the particles had been moved to the sieve plate, where they aggregated (Figure 6D). In places, the particles were held together by strandlike material that made an elastic and sticky impression (Figure 6C).
To approximate the rate of irradiation that provokes detachment and clogging of the parietal proteins (Figure 6A to D), laser energy flow rates were measured in the focus plane of the objectives. With an image size of 625 x 625 µm and at a laser energy flow rate of 600 µW, which is calculated to represent a laser energy fluence rate of ~1500 W m-2, no effect on the translocation capacity of the SEs was observed for hours (Figure 4A and Figure 4B). It should be noted that the calculations overestimate the energy fluence rates seen by the tissues, because medium and overlying tissues cause loss of intensity. A time scan of ~10 min (Figure 4F), at an energy flow rate of 45 µW representing 7000 W m-2 at an image size of 80 x 80 µm, did not affect the ultrastructure of the SEs as well. Immediate effects were visible after raising the energy fluence rate to ~30,000 W m-2 (energy flow rate of 100 µW, with an image size of 61.2 x 61.2 µm; Figure 6A and Figure 6B).
We wanted to determine whether the sedimentary layer occasionally observed in actually translocating SEs (Figure 4F and Figure 5B) is an "in-borne" artifact of laser radiation. We believe that it is not for a number of reasons. (1) Under low laser intensity, proteinaceous sediment in intact SEs resided, if present at all, at either side of the sieve plate (Figure 4F and Figure 5B). By contrast, sedimentation in SEs disturbed by intense laser irradiation always occurred at the side of the sieve plate facing the direction of the translocation stream (Figure 6D). (2) Sediment, originally present at the other side of the sieve plate, was dislodged by intense laser irradiation and was swept away by the mass flow to the next sieve plate (Figure 6A and Figure 6C). After detachment of the material, the sieve pores remained behind for awhile as empty spaces (Figure 6B). As a result, the detached material accumulated against the sieve plates in laser-irradiated SEs, whereas in unaffected SEs the sediment stays at all margins of the cell. (3) In phloem areas preexposed to intense laser light, translocation of phloem-mobile dyes was never observed, whereas fluorochrome movement was unhampered in the untreated adjacent regions (results not shown). This suggests a retardation of fluorochrome movement by the sieve plate-associated sediments in SEs impaired by intense radiation.
Effects of Mechanical Injury on the SE Structure
Remarkably, the P plastids stayed unaffected by intensified laser radiation (Figure 6A to D). The responses to injury by laser light and to mechanical damage may differ, also because of the absence of P plastids in mechanically damaged tissues (Figure 1D and Figure 1E). To identify the processes involved in the response to mechanical damage, originally intact SEs were exposed to rate-controlled mechanical damage. This study comprised observations on the effects of impalement with microelectrode tips of various diameters on SE structure (Figure 7 Figure 8 Figure 9).
|
|
|
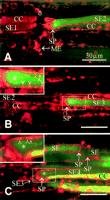
In intact SEs, the P plastids were evenly distributed and located against the cell wall (Figure 7A and Figure 8A). As soon as a microelectrode (diameter of ~1 µm) was introduced into an intact SE, the pressure was instantly released in the impaled SE. The pressure release was indicated by bending of the sieve plate immediately after impalement (Figure 7B). Simultaneously, all P plastids in the damaged SE and most P plastids in the adjoining SE exploded, leaving their fluorescent coats on the wall (Figure 7B). Logically, the burst of the P plastids was ascribed to the abrupt loss of pressure inside of the SE. Combined detection of CDCFDA and RH-160 in the same SE (Figure 7C and Figure 7D) revealed that penetration of the needle provoked the formation of a CDCFDA-tagged plug on the sieve plate within 1 min (Figure 7C). In the course of up to 5 min, the fluorescence of the plug intensified (Figure 7D), and additional fluorescent material of a different nature traveled to the plug (Figure 7D), giving rise to a composed sedimentary plug similar to the one on the sieve plates in tissue slices (Figure 1D). The question is whether the plug gradually condensed or whether the staining gradually intensified during the period of observation.
The harsh penetration of the microelectrode impeded sealing of the plasma membrane so that the fluorescent cytosolic content leaked out of the SE (Figure 7D). Apparently, the plug produced in the adjacent SE partly occluded the sieve plate, because some fluorescent material stayed within this SE (Figure 7D).
That the pressure release, and not a "wound reaction," provokes the burst of P plastids was demonstrated by careful insertion of a microelectrode with a minuscule electrode tip (diameter of ~0.1 µm). Insertion of such a microelectrode tip (Figure 8A) was not expected to cause any pressure release from the SE, provided that the plasma membrane sealing was perfect. Expectedly, the turgor pressure of the SE was insufficient to overcome the capillary forces inside of a tip of this size. Actually, P plastids remained intact during and after impalement into RH-160/HPTSA-stained intact SEs and no plug was formed (Figure 8A). As soon as the electrode was retracted, pressure appeared to be released through the small hole in the plasma membrane left behind by the electrode tip. The P plastids immediately exploded after withdrawal of the electrode, and the onset of a plug formation became visible (Figure 8B). The minor turgor loss had only local effects: the P plastids in the next SE remained intact (Figure 8B). The plugs provoked by mechanical damage did not bind to RH-160 (Figure 7D and Figure 8B) in contrast to the sediment caused by laser irradiation (Figure 4F, Figure 5B, and Figure 6D).
After severe damage made by the penetration of a coarse microelectrode tip (diameter of ~10 µm), a flexible and gradual response to injury was inferred from the behavior of four successive SEs (Figure 9A to C). The impaled SE (SE1) lost all CDCFDA, indicative of the loss of the cytosolic content. In the next SE (SE2), meanwhile, a plug had been built up that partly prevented fluorochrome leakage (Figure 9A). Before plugging the sieve plate, part of the SE content must have been lost. Fluorescence intensity of SE2 was appreciably lower than that of SE3; it was so much lower that the presence of CDCFDA in SE2 became invisible at a laser intensity sufficient to visualize CDCFDA in SE3 (Figure 9A and Figure 9B). Moreover, sieve plate plugging was less massive, and some P plastids remained intact in SE3 (cf. Figure 9A and Figure 9B). Only a minor plug had been formed in SE4, whereas most P plastids were still intact (Figure 9C).
|
|
DISCUSSION |
|---|
Identification and Localization of the Structural Components of Intact SEs
The phloem-mobile fluorochromes (CFDA, CDCFDA, and HPTSA) barely stained the structural components of intact SEs. The exceptions were CFDA labeling the starch grains in the P plastids (Figure 4C and Figure 4D) and HPTSA labeling faintly large spindle-shaped bodies of proteinaceous material (Figure 5B). These bodies are likely identical to the massive P protein clusters, which are typical of Fabaceae. Electron microscopy of soybean (![]()
![]()
![]()
![]()
Highly informative on the SE structure was the local application of RH-160, which originally was meant to visualize only the plasma membrane. The high resolution of the objective and the distinct fluorescence of the plasma membrane lining allowed us to observe the plasma membrane traversing the sieve plate pores and the pore plasmodesma units (Figure 4E).
Given the membranous nature of the P plastid coats, binding of RH-160 is logical on the assumption that RH-160 is slowly membrane permeant. As suspected by many investigators but never shown convincingly by using electron microscopy, P plastids are no doubt evenly distributed over the plasma membrane in intact SEs (Figure 4F, Figure 5B, Figure 6A, Figure 8A, and Figure 10). A unique reconstruction from light microscopic images of sections of Aristolochia phloem (![]()
|
The assessment of the other parietal structures stained by RH-160 is difficult, because the initial assumption that RH-160 binds only to membranes does not seem to be true. The strands that emerge from the parietal proteins in mechanically damaged SEs (Figure 1E and Figure 1F) are composed of nonmembranous material and yet stained by RH-160. The fluorescent material likely represents ER labyrinths or parietal proteins. Convoluted ER or appressed stacks of ER are abundant in the SEs of some species (![]()
![]()
![]()
![]()
![]()
Structure and Anchoring of P Plastids
P plastids are long-established organelles in the SEs of dicotyledons, the substructure of which is strongly species specific (![]()
![]()
![]()
![]()
Our work seems to shed some light on the function of the P plastids. The injury experiments with microelectrodes of various diameters (Figure 6 Figure 7 Figure 8 Figure 9) demonstrate that a pressure drop in the SE cytosol triggers the disruption of the P plastids. In particular, the careful insertion and quick retraction of fine microelectrode tips (Figure 8) suggest that pressure release rather than the injury acts as the primary signal for wounding.
After explosion of the P plastids, the membranous fluorescent coats were still visible, attached to the wall, whereas the starch granules were recognizable near the sieve plate as particles faintly stained by CFDA (Figure 2). The liberated cores of starch were the particles often seen by light microscopy while making Brownian motion near the sieve plates of severed SEs (![]()
![]()
The consistent parietal location of the P plastids and their remnants indicates that the P plastids must be anchored in some way (Figure 4F). The attachment must be stable; time scans did not show any movement of the P plastids, and the basal plastid remnants did not displace after bursting (Figure 6A to D and Figure 7B to D). Electron microscopy actually showed P plastids attached to ER, which in turn may be continuous with the plasma membrane (![]()
![]()
Morphology and Definition of the Proteinaceous Compounds of the SEs
Location, form, and chemical nature of the proteins are very variable and obviously species specific. The complexity has been increased by the various degrees of damage response due to different preparation and fixation procedures. That several soluble proteins may turn into an unsoluble form in response to injury further complicates the identification of the structures. The damage responses in our study can be assessed only when the structural proteins involved are defined properly.
At least 150 mainly low molecular weight proteins occur in the SEs (![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Sieve Pores: Open or Plugged?
![]()
The restriction of the phloem-mobile probes to the SE/CC complexes confirms once again that the few plasmodesmata between the CCs and the phloem parenchyma (![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Response to Laser Light
Strong laser irradiation provokes detachment of the RH-160 reactive parietal sediment. RH-160 is reported to be a membrane marker and therefore would indicate detachment of stacked ER, which is abundantly present in the SEs of fava bean (I. Dörr, personal communication). Yet, we believe that the major part of the detached particles are parietal proteins for several reasons. (1) Thick and irregular packs of similar material were shown to be proteinaceous in several species (e.g., ![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
The response to laser light intensities (Figure 6) likely does not reflect the behavioral reaction to natural light conditions. The intensities exceed by far the solar constant of 1400 W m-2. The reaction of the SEs may be due to a rise in temperature imposed by the incident laser light and may express a defense mechanism against high temperatures. Incidently, the results warn against unlimited laser radiation for the study of living cells.
Response to Mechanical Damage
The reactions to mechanical injury and to laser light are clearly different. However, both may occur simultaneously, in particular when the damage is severe (Figure 2). The main difference between the reactions lies in the bursting of the P plastids. The disruption of the P plastids is due to the pressure release in the SEs and not to immediate wounding responses (Figure 8A and Figure 8B). That the P plastids set free coagulating substances is the most plausible explanation for the massive plug formation on the sieve plates, given the time frame and the order of events (Figure 7 Figure 8 Figure 9). In an earlier study in which P plastids were burst by iodine treatment, the liberated starch grains were hypothesized to play a role in sieve plate plugging (![]()
Most likely, the crystalline P protein body represents the nucleus of the plug formation in fava bean. The putative scenario of plug formation implies that dispersion of the P protein bodies should go along with breakage of the P plastids. Such a coincidence was confirmed by electron microscopic images of soybean (![]()
![]()
![]()
Apart from the coagulates from P protein material and P plastid content, the plugs also are composed of material from the parietal protein. This material was visible as a more intensely labeled layer on the primary plug (Figure 1A to D) and was connected by robust strands with the parietal plasma membrane regions. The strands that emerged in severely damaged SEs (Figure 1E and Figure 1F) were also seen as bundles of P protein filaments in electron micrographs (![]()
![]()
Efficiency of the Plugging Reactions
In conclusion, P plastids explode in immediate response to pressure release in the SEs and set substances free that react with the P protein body material. The product is a conglomerate of substances that plug the sieve pores. Activated by the surge produced by the pressure release, parietal proteins are torn away or detach from the SE plasma membrane, building a cover layer on the developing plug (Figure 2). The elasticity and stickiness of the parietal material give rise to an irregular system of strands. The vigor of the pressure disturbance determines the morphology of the strand system (Figure 2).
The presence of parietal P plastids and ER (Figure 10) did not prevent mass flow in intact SEs (Figure 4F, Figure 5A, and Figure 5B). Hence, the deposition of parietal proteins on the sieve plates (Figure 6A to D) expectedly only retards solute passage through the sieve plate pores to some degree. When fluorescent dyes were injected into the SEs, depositions occurred, probably in response to the impalement of the microelectrode (![]()
![]()
![]()
![]()
![]()
![]()
![]()
|
|
METHODS |
|---|
Plant Material
Pot plants of Vicia fava cv Witkiem major (Nunhems Zaden BV, Haelen, The Netherlands) were grown in a greenhouse under standard conditions of 20°C at 60 to 70% relative humidity during a 14-hr-day and 10-hr-dark period, with daylight plus additional lamp light (model SONT Agro 400 W; Philips, Eindhoven, The Netherlands) to a minimum irradiance of 250 µmol m-2 sec-1 at plant level. Plants were used for experiments 17 to 21 days after germination.
Preparation for Observation by Confocal Laser Scanning Microscopy
A few cortical cell layers were locally removed down to the phloem from the lower side of the main vein of a mature leaf. The layers were removed by manual paradermal slicing with a new Wilkinson razor blade (Figure 3, position 1), while avoiding damage of the phloem. Immediately after cutting a shallow window (~10 mm long and 2 mm wide), we bathed the free-lying phloem tissue in a medium with 10 mol m-3 KCl, 10 mol m-3 CaCl2, and 5 mol m-3 NaCl in double-distilled water. To allow the cells to create their own apoplasmic environment, the medium was not buffered. The leaf was fixed in an upside-down position with two-side adhesive tape on a convex plastic slide. The state of the tissue was checked microscopically. If the phloem appeared to be undamaged, a second window (10 x 2 mm), 2 to 3 cm acropetal from the first one, was manufactured (Figure 3, position 2) and bathed in the unbuffered medium mentioned previously. The convex slide carrying the leaf, still attached to the whole plant, was mounted on the stage of a confocal laser scanning microscope. The tissue windows were blotted just before application of fluorochrome solutions.
Fluorescent Probes
To be used as phloem mobile tracers, 10 mg of 5(6)-carboxyfluorescein diacetate (CFDA; C-195) and 5(6)-carboxy-2',7'-dichlorofluorescein diacetate (CDCFDA; C-369) were each dissolved in a minimum volume of 500 mol m-3 KOH. The stock was diluted in double-distilled water up to 20 mL. KCl, CaCl2, and NaCl were added to final concentrations of 10, 10, and 5 mol m-3, respectively. The pH was adjusted to 6.3 with 1000 mol m-3 HCl.
As a phloem-mobile compound alternative to the CFDA derivatives, 8-acetoxypyrene-1,3,6-trisulfonic acid trisodium salt (HPTSA; A-396) was used, which was dissolved in double-distilled water to a final concentration of 20 mg mL-1. KCl, CaCl2, and NaCl were added to final concentrations of 10, 10, and 5 mol m-3, respectively.
Lucifer yellow CH, potassium salt (LYCH; L-1177) was dissolved in double-distilled water to a concentration of 7% (w/v). The dye was kept in darkness at 4°C for ~2 weeks, during which the unsolubilized crystals sedimented. The supernatant was used for filling microelectrode tips. Sulforhodamin G (Aldrich, Milwaukee, WI) was dissolved in 0.1 mol m-3 NaOH-Mes buffer, pH 6, to a final concentration of 0.01% and used for filling microelectrode tips.
For its use as a membrane stain, 25 mg of N-(4-sulfobutyl)-4-(4-(4-dibutylamino)phenyl)butadienyl)pyridinium inner salt (RH-160, S-1107) was solubilized in 1 mL of ethanol. Two microliters of the stock was added directly to the bathing medium. Apart from sulforhodamin G, all fluorochromes were purchased from Molecular Probes (Eugene, OR; the codes are from the 1996 catalog).
Microscopy
Phloem tissue was examined with a Leica (Heidelberg, Germany) TCS 4D confocal laser scanning microscope. CFDA, CDCFDA, LYCH, and sulforhodamin G were excited by the 488-nm line produced by a 75-mW argon/krypton laser (Omnichrome, Chino, CA). The emission was observed by using a fluorescein band pass filter.
HPTSA was excited by the 488- and 476-nm lines of the argon/krypton laser. The emitted light was split by a 580-nm beam splitter, and short wavelengths were observed by using a 510-nm long-pass filter. RH-160 was excited by the 564-nm line of the 75-mW argon/krypton laser, and a 590-nm long pass filter was used for observation.
For two-channel scans with CFDA, CDCFDA, HPTSA, LYCH, or sulforhodamin G, on the one hand, and RH-160, on the other, the 488- and the 564-nm lines of the laser were used simultaneously for excitation. A 580-nm beam splitter passed the light to the two detector heads, and the appropriate emission filters were selected for observation. The observations were conducted with a PL Fluotar x16/0.50 IMM objective (Leica; Figure 4A and Figure 4B) or a PL APO x63/1.20 W CORR objective (Leica; Figure 1, Figure 4C to F, Figure 5 Figure 6 Figure 7, and Figure 9).
Observation of the Tissues
In initial experiments, petiole slices were prepared in which phloem tissue was damaged to examine the effects of severe injury. The damaged tissue was stained with CDCFDA or RH-160. In subsequent experiments with intact plants, the apical window served the application of phloem-mobile fluorochromes, the basal one the administration of RH-160 and the observation of the phloem tissue. The movement of phloem-mobile fluorochromes was observed from ~30 min after fluorochrome application at the apical window. The working distance of 220 µm between objective and object left a space just large enough to insert microelectrode tips. The microelectrodes were advanced by a four-axis joystick micromanipulator (model MMW-204; Narishige, Tokyo, Japan) that was installed on the stage of the confocal laser scanning microscope in the upright configuration. The upright position enabled observation of intact plants and the fixed micromanipulator continuous focusing during the manipulation. The minute working distance required electrodes with a long strong shank and rapidly tapering tip. Microelectrodes with a fine (diameter of ~0.1 µm) and medium (diameter of ~1 µm) tip size were pulled from borosilicate glass capillaries (1 mm outer diameter, 0.58 mm inner diameter with an inner filament; Clark Electromedical Instruments, Reading, UK) with a laser puller (model P-2000; Sutter Instruments, Novato, CA). The microelectrodes with a coarse tip (diameter of ~10 µm) were manufactured by breaking the tip from 1-µm microelectrodes.
Measurement of Laser Energy Flow Rates
Laser energy flow rates and energy fluence rates (see ![]()
|
|
ACKNOWLEDGMENTS |
|---|
All original images and a few extra time scans (movies) related to this article can be retrieved from the following internet page: http:/www. uni-giessen.de/ ˜gf1114/rgvbel/index.htm. We extend our thanks to Denton Prior and Dr. Karl Oparka (Scottish Crop Research Institute, Invergowrie, Dundee, Scotland) for all forms of support during the confocal laser scanning microscope installation, Dr. Hubert Felle (University of Giessen) for assistance in fabrication of the microelectrodes, Guido Steude (University of Giessen) for measuring laser energy flow rates, Dr. Inge Dörr (University of Kiel, Germany) for providing us with unpublished work, Volkmar Thier and Sven Tränkner (Senckenbergische Naturforschende Gesellschaft, Frankfurt, Germany) for computer processing of the confocal laser scanning microscope images, Dr. H.-Dietmar Behnke (University of Heidelberg, Germany) and Dr. Alexander Schulz (University of Kiel) for helpful criticism, and Manfred Jung, Peter Schneider, and Dorothee Schroth (Leica, Bensheim, Germany) for continuous advice and support.
Received July 3, 1997; accepted November 19, 1997.
|
|
REFERENCES |
|---|
Barclay, G.F., Oparka, K.J., and Johnson, R.P.C. (1977) Induced disruption of sieve element plastids in Heracleum mantagazzianum L. J. Exp. Bot. 28:709-717[ISI]
Behnke, H.D. (1981) Sieve-element characters. Nord. J. Bot. 1:381-400
Behnke, H.D., and Sjolund, R.D. (1990). Sieve Elements. (Berlin: Springer-Verlag).
Canny, M.J.P. (1975). Mass transfer. In Encyclopedia of Plant Physiology, Vol. 1, M.H. Zimmermann and J.A. Milburn, eds (Berlin: Springer-Verlag), pp. 139-153.
Cronshaw, J., and Sabnis, D.D. (1990). Phloem proteins. In Sieve Elements, H.D. Behnke and R.D. Sjolund, eds (Berlin: Springer-Verlag), pp. 257-283.
Esau, K. (1969). The Phloem. Encyclopedia of Plant Anatomy, Vol. 5 (Berlin: Borntraeger).
Esau, K. (1978) Developmental features of the primary phloem in Phaseolus vulgaris L. Ann. Bot. 42:1-13
Esau, K., and Cronshaw, J. (1967) Tubular components in cells of healthy and tobacco mosaic virus-infected Nicotiana.. Virology 33:26-35[ISI][Medline]
Esau, K., and Gill, R.H. (1972) Nucleus and endoplasmic reticulum in differentiating root protophloem of Nicotiana tabacum.. J. Ultrastruct. Res. 41:160-175[ISI][Medline]
Evert, R.F. (1990). Dicotyledons. In Sieve Elements, H.D. Behnke and R.D. Sjolund, eds (Berlin: Springer-Verlag), pp. 103-137.
Evert, R.F., and Derr, W.F. (1964) Callose substance in sieve elements. Am. J. Bot. 51:552-559[ISI]
Evert, R.F., Tucker, C.M., Davis, J.D., and Deshpande, B.P. (1969) Light microscope investigation of sieve element ontogeny and structure in Ulmus americana.. Am. J. Bot. 56:999-1017[ISI]
Evert, R.F., Eschrich, W., and Eichhorn, S.E. (1973) P protein distribution in mature sieve elements of Cucurbita maxima.. Planta 109:193-210[ISI]
Fensom, D.S. (1957) The bioelectric potentials of plants and their functional significance. I. An electrokinetic theory of transport. Can. J. Bot. 35:573-582
Fisher, D.B. (1975) Structure of functional soybean sieve elements. Plant Physiol. 56:555-569[ISI]
Fisher, D.B., Wu, Y., and Ku, M.S.B. (1992) Turnover of soluble proteins in the wheat sieve tube. Plant Physiol. 100:1433-1441[ISI]
Giaquinta, R.T., and Geiger, D.R. (1973) Mechanism of inhibition of translocation by localized chilling. Plant Physiol. 51:372-377[ISI]
Goodall, H., and Johnson, M.H. (1982) Use of carboxyfluorescein diacetate to study formation of permeable channels between mouse blastomers. Nature 295:524-526[ISI][Medline]
Grignon, N., Touraine, B., and Durand, M. (1989) 6(5)Carboxyfluorescein as a tracer of phloem sap translocation. Am. J. Bot. 76:871-877[ISI]
Hartig, T. (1837) Vergleichende Untersuchungen über die Organisation des Stammes der einheimischen Waldbäume. Jahresber. Fortschr. Forstwiss. Forstl. Naturkd. 1:125-168
Hartig, T. (1854) Über die Querscheidewände zwischen den einzelnen Gliedern der Siebröhren in Cucurbita pepo.. Bot. Z. 12:51-54
Hartig, T. (1860) Beiträge zur physiologischen Forstbotanik. Allgemeine Forst- u. Jagdzeitschrift 36:257-261
Hayes, P.M., Patrick, J.W., and Offler, C.E. (1987) The cellular pathway of radial transfer of photosynthates in stems of Phaseolus vulgaris L.: Effects of cellular plasmolysis and p-chloromercuribenzene sulphonic acid. Ann. Bot. 59:635-642
Johnson, R.P.C., Freundlich, A., and Barclay, G.F. (1976) Transcellular strands in sieve tubes; What are they? J. Exp. Bot. 101:1117-1136
Kempers, R., and van Bel, A.J.E. (1997) Symplasmic connections between sieve element and companion cell in the stem of Vicia fava L. have a molecular exclusion limit of at least 10 kD. Planta 201:195-201[ISI]
Kempers, R., Ammerlaan, A., and van Bel, A.J.E. (1998) Symplasmic
constriction and ultrastructural features of the sieve element/companion
cell complex in the transport phloem of apoplasmatically and symplasmatically
phloem loading species. Plant Physiol. 116:271-278
Kollmann, R. (1973). Cytologie des phloems. In Grundlagen der Cytologie, G.C. Hirsch, H. Ruska, and P. Sitte, eds (Jena, Germany: Fischer), pp. 479-505.
Lawton, D.M. (1978a) P protein crystals do not disperse in uninjured sieve elements in roots of runner bean (Phaseolus multiflorus) fixed with glutaraldehyde. Ann. Bot. 42:353-361
Lawton, D.M. (1978b) Ultrastructural comparison of the tailed and tailless P protein crystals respectively of runner bean (Phaseolus vulgaris) and garden pea (Pisum sativum) with tilting stage electron microscopy. Protoplasma 97:1-11[ISI]
Münch, E. (1930). Die Stoffbewegung in der Pflanze. (Jena, Germany: Fischer).
Nakamura, S., Hayashi, H., Mori, S., and Chino, M. (1993) Protein phosphorylation in the sieve tubes of rice plants. Plant Cell Physiol. 34:927-933[ISI]
Oparka, K.J., and Johnson, R.P.C. (1978) Endoplasmic reticulum and crystalline fibrils in the root protophloem of Nymphoides peltata.. Planta 143:21-27[ISI]
Oparka, K.J., Viola, R., Wright, K.M., and Prior, D.A.M. (1992). Sugar transport and metabolism in potato tuber. In Carbon Partitioning within and between Organisms, C.J. Pollock, J.F. Farrar, and A.J. Gordon, eds (Oxford, UK: BIOS), pp. 91-114.
Oparka, K.J., Duckett, C.M., Prior, D.A.M., and Fisher, D.B. (1994) Real-time imaging of phloem unloading in the root tip of Arabidopsis.. Plant J. 6:759-766[CrossRef][ISI]
Palevitz, B.A., and Newcomb, E.H. (1971) The ultrastructure and development of tubular and crystalline P protein in the sieve elements of certain papilionaceous legumes. Protoplasma 72:399-426[ISI]
Patrick, J.W., and Offler, C.E. (1996) Post-sieve element transport of photoassimilates in sink regions. J. Exp. Bot. 47:1165-1177[Abstract]
Rhodes, J.D., Thain, J.F., and Wildon, D.C. (1996) The pathway for systemic electrical signal conduction in the wounded tomato plant. Planta 200:50-57[ISI]
Roberts, C.J., Raymond, C.K., Yamashiro, C.T., and Stevens, T.H. (1991) Methods to studying the yeast vacuole. Methods Enzymol. 194:644-661[ISI][Medline]
Robidoux, J., Sandborn, E.B., Fensom, D.S., and Cameron, M.L. (1973) Plasmatic filaments and particles in mature sieve elements of Heracleum sphondylium under the electron microscope. J. Exp. Bot. 79:349-359
Salisbury, F.B. (1996). Units, Symbols, and Terminology for Plant Physiology. (Oxford, UK: Oxford University Press).
Spanner, D.C. (1958) The translocation of sugar in sieve tubes. J. Exp. Bot. 9:332-342[ISI]
Spanner, D.C. (1970) The electroosmotic theory of phloem transport in the light of recent measurements of Heracleum phloem. J. Exp. Bot. 21:325-334[ISI]
Spanner, D.C. (1978) Sieve-plate pores, open or occluded? A critical review. Plant Cell Environ. 1:7-20
Thaine, R. (1961) Transcellular strands and particle movement in mature sieve tubes. Nature 192:772-773[ISI]
Thaine, R. (1962) A translocation hypothesis based on the structure of plant cytoplasm. J. Exp. Bot. 13:152-160[ISI]
Thaine, R. (1969) Movement of sugars through plants by cytoplasmic pumping. Nature 222:873-875[ISI]
Thorsch, J., and Esau, K. (1981a) Changes in the endoplasmic reticulum during differentiation of a sieve element in Gossypium hirsutum.. J. Ultrastruct. Res. 74:183-194[ISI][Medline]
Thorsch, J., and Esau, K. (1981b) Nuclear degeneration and the association of endoplasmic reticulum with the nuclear envelope and microtubules in maturing sieve elements of Gossypium hirsutum.. J. Ultrastruct. Res. 74:195-204[ISI][Medline]
Toth, P.T., Bindokas, V.P., Bleakman, D., Colmers, W.F., and Miller, R.J. (1993) Mechanism of presynaptic inhibition by neuropeptide Y at sympathetic nerve terminals. Nature 364:635-638[CrossRef][ISI][Medline]
van Bel, A.J.E. (1995). The low profile directors of carbon and nitrogen economy in plants: Parenchyma cells associated with translocation channels. In Plant Stems: Physiology and Functional Morphology, B.L. Gartner, ed (San Diego, CA: Academic Press), pp. 205-222.
van Bel, A.J.E., and Kempers, R. (1991) Symplastic isolation of the sieve element-companion cell complex in the phloem of Ricinus communis and Salix alba stems. Planta 183:69-76[ISI]
van Bel, A.J.E., and van Rijen, H.V.M. (1994) Microelectrode-recorded development of the symplasmic autonomy of the sieve element/companion cell complex in the stem phloem of Lupinus luteus L. Planta 192:165-175[ISI]
van der Schoot, C., and van Bel, A.J.E. (1989) Glass microelectrode measurements of sieve tube membrane potentials in internode discs and petiole strips of tomato (Solanum lycopersicum L.). Protoplasma 149:144-154[ISI]
van der Schoot, C., and van Bel, A.J.E. (1990) Mapping membrane potentials and dye coupling in internodal tissues of tomato (Solanum lycopersicum L.). Planta 182:9-21[ISI]
Walsh, M.A., and Popovich, T.M. (1977) Some ultrastructural aspects of metaphloem sieve elements in the aerial stem of the holoparasitic angiosperm Epifagus virginiana (Orobranchaceae). Am. J. Bot. 64:326-336[ISI]
Wang, H.L., Offler, C.E., Patrick, J.W., and Ugalde, T.D. (1994) The cellular pathway of photosynthate transfer in the developing wheat grain. I. Delineation of a potential transfer pathway using fluorescent dyes. Plant Cell Environ. 17:257-266[ISI]
Wang, N., and Fisher, D.B. (1994) The use of fluorescent tracers to
characterize the post-phloem transport pathway in maternal tissue of developing
wheat grains. Plant Physiol. 104:17-27
Wergin, W.P., and Newcomb, E.H. (1970) Formation and dispersal of crystalline P protein in sieve elements of soybean (Glycine max L.). Protoplasma 71:365-388[ISI]
Wright, K.M., and Oparka, K.J. (1996) The fluorescent probe HPTS as a phloem-mobile, symplastic tracer: An evaluation using confocal laser scanning microscopy. J. Exp. Bot. 47:439-445[Abstract]
Yin, Z.H., Neimanis, S., Wagner, U., and Heber, U. (1990) Light-dependent pH changes in leaves of C3 plants. Planta 182:244-252[ISI]
This article has been cited by other articles:
 |
G.-L. Wu, X.-Y. Zhang, L.-Y. Zhang, Q.-H. Pan, Y.-Y. Shen, and D.-P. Zhang Phloem Unloading in Developing Walnut Fruit is Symplasmic in the Seed Pericarp and Apoplasmic in the Fleshy Pericarp Plant Cell Physiol., October 15, 2004; 45(10): 1461 - 1470. [Abstract] [Full Text] [PDF] |
||||
 |
B.-C. Yoo, F. Kragler, E. Varkonyi-Gasic, V. Haywood, S. Archer-Evans, Y. M. Lee, T. J. Lough, and W. J. Lucas A Systemic Small RNA Signaling System in Plants PLANT CELL, August 1, 2004; 16(8): 1979 - 2000. [Abstract] [Full Text] [PDF] |
||||
 |
L.-Y. Zhang, Y.-B. Peng, S. Pelleschi-Travier, Y. Fan, Y.-F. Lu, Y.-M. Lu, X.-P. Gao, Y.-Y. Shen, S. Delrot, and D.-P. Zhang Evidence for Apoplasmic Phloem Unloading in Developing Apple Fruit Plant Physiology, May 1, 2004; 135(1): 574 - 586. [Abstract] [Full Text] [PDF] |
||||
|
|
D. A. Bird, V. R. Franceschi, and P. J. Facchini A Tale of Three Cell Types: Alkaloid Biosynthesis Is Localized to Sieve Elements in Opium Poppy PLANT CELL, November 1, 2003; 15(11): 2626 - 2635. [Abstract] [Full Text] [PDF] |
||||
|
|
S. Dinant, A. M. Clark, Y. Zhu, F. Vilaine, J.-C. Palauqui, C. Kusiak, and G. A. Thompson Diversity of the Superfamily of Phloem Lectins (Phloem Protein 2) in Angiosperms Plant Physiology, January 1, 2003; 131(1): 114 - 128. [Abstract] [Full Text] [PDF] |
||||
 |
A. I. Prokhnevsky, V. V. Peremyslov, A. J. Napuli, and V. V. Dolja Interaction between Long-Distance Transport Factor and Hsp70-Related Movement Protein of Beet Yellows Virus J. Virol., October 2, 2002; 76(21): 11003 - 11011. [Abstract] [Full Text] [PDF] |
||||
|
|
A. Itaya, F. Ma, Y. Qi, Y. Matsuda, Y. Zhu, G. Liang, and B. Ding Plasmodesma-Mediated Selective Protein Traffic between "Symplasmically Isolated" Cells Probed by a Viral Movement Protein PLANT CELL, September 1, 2002; 14(9): 2071 - 2083. [Abstract] [Full Text] [PDF] |
||||
 |
B.-C. Yoo, J.-Y. Lee, and W. J. Lucas Analysis of the Complexity of Protein Kinases within the Phloem Sieve Tube System. CHARACTERIZATION OF CUCURBITA MAXIMA CALMODULIN-LIKE DOMAIN PROTEIN KINASE 1 J. Biol. Chem., May 3, 2002; 277(18): 15325 - 15332. [Abstract] [Full Text] [PDF] |
||||
|
|
N. Rouhier, E. Gelhaye, P.-E. Sautiere, A. Brun, P. Laurent, D. Tagu, J. Gerard, E. de Fay, Y. Meyer, and J.-P. Jacquot Isolation and Characterization of a New Peroxiredoxin from Poplar Sieve Tubes That Uses Either Glutaredoxin or Thioredoxin as a Proton Donor Plant Physiology, November 1, 2001; 127(3): 1299 - 1309. [Abstract] [Full Text] [PDF] |
||||
|
|
N. A. Eckardt A Calcium-Regulated Gatekeeper in Phloem Sieve Tubes PLANT CELL, May 1, 2001; 13(5): 989 - 992. [Full Text] |
||||
|
|
M. Knoblauch, W. S. Peters, K. Ehlers, and A. J. E. van Bel Reversible Calcium-Regulated Stopcocks in Legume Sieve Tubes PLANT CELL, May 1, 2001; 13(5): 1221 - 1230. [Abstract] [Full Text] |
||||
 |
S. Haupt, K. J. Oparka, N. Sauer, and S. Neumann Macromolecular trafficking between Nicotiana tabacum and the holoparasite Cuscuta reflexa J. Exp. Bot., January 1, 2001; 52(354): 173 - 177. [Abstract] [Full Text] |
||||
|
|
K. J. Oparka and R. Turgeon Sieve Elements and Companion Cells—Traffic Control Centers of the Phloem PLANT CELL, April 1, 1999; 11(4): 739 - 750. [Full Text] |
||||
|
|
B. Golecki, A. Schulz, and G. A. Thompson Translocation of Structural P Proteins in the Phloem PLANT CELL, January 1, 1999; 11(1): 127 - 140. [Abstract] [Full Text] |
||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
| ASPB JOURNALS | THE PLANT CELL | PLANT PHYSIOLOGY | |
|---|---|---|---|