{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
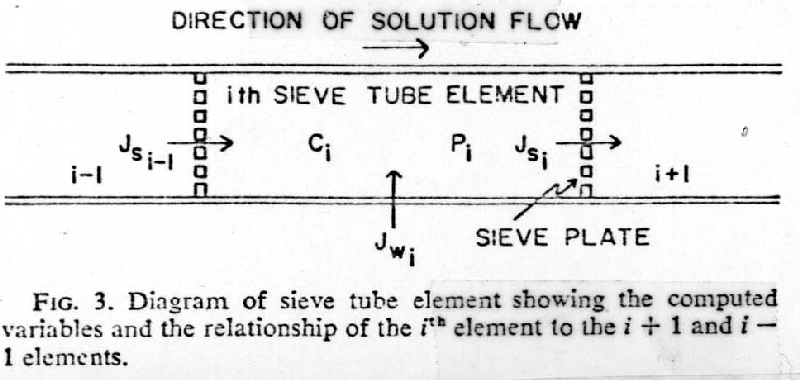
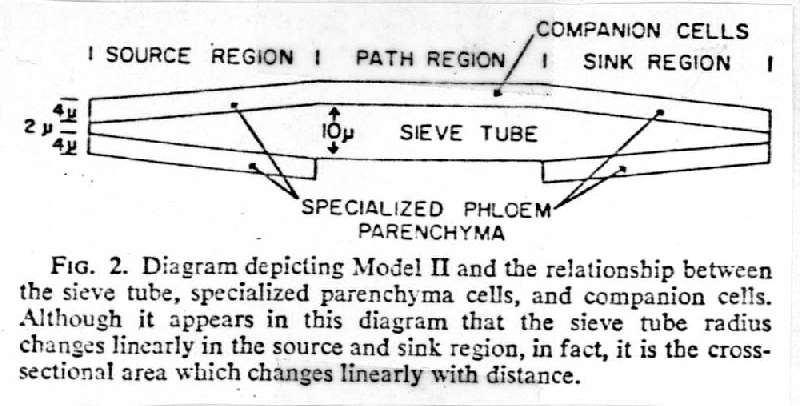
I. Anatomical variations within the Phloem
A. Conducting components
1. Generally occupy 20% of total cross sectional area of Phloem tissue
a.
About a 10 fold difference in radius within a plant
b. Largest in secondary phloem, smallest in minor leaf veins
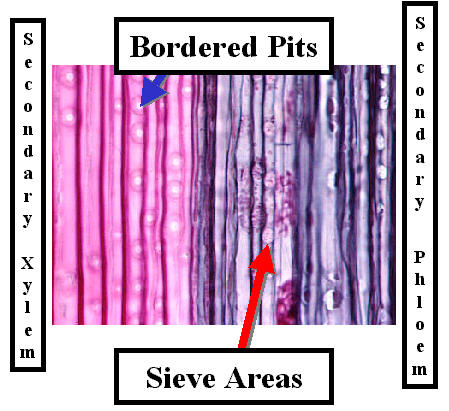
2. Sieve tube members form sieve tubes
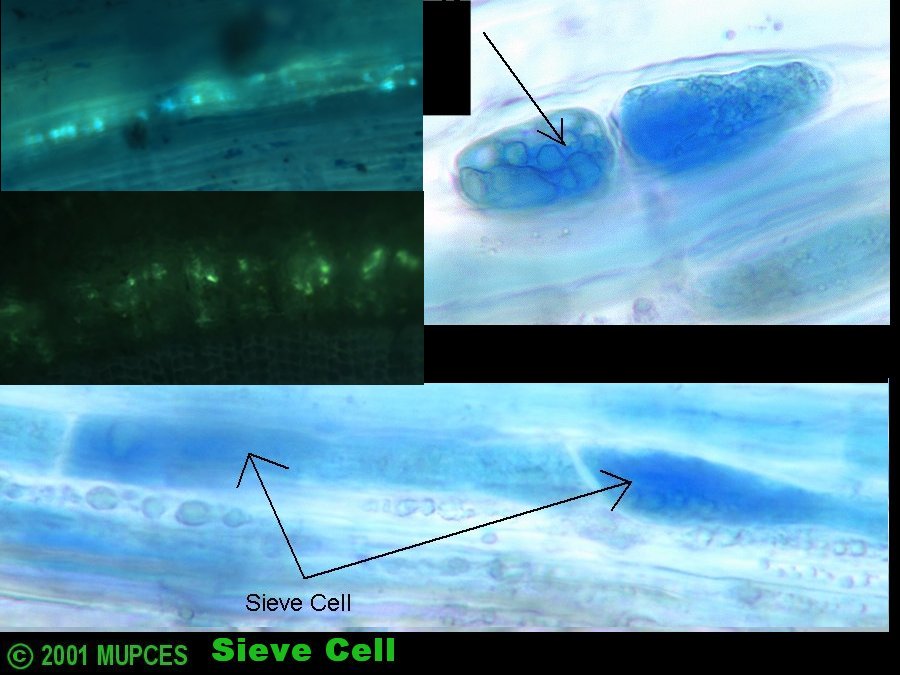
3. Sieve cells
4. Sieve pores/areas usually 1.0 um in diameter
5. Amount of callose under physiological control
a. Definitive = small amount found in functional sieve elements
b. Wound = amplified amount produced in response to wounds
c. Dormancy = amplified amount produced during onset of dormancy,
decreases when growth resumes
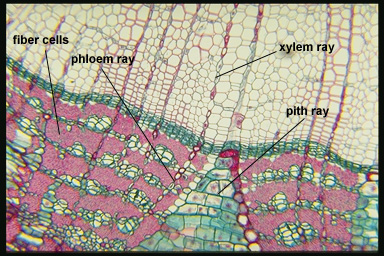
1. Parenchyma
a. Axial
b. Ray, often dilated
2. Companion Cells or Albuminous Cells
3. Transfer Cells
C. Structural components
1. Bast fibers
2. Sclereids
D. First formed phloem (Protophloem) typically obliterated after Metaphloem formation
1. Sieve tubes of monocot protophloem not associated with companion cells
2. Metaphloem sieve tube elements
a. Longer lived
b. Usually longer and wider
c. More distinct sieve areas
d. Literature states that no fibers develop in Metaphloem
E. Metaphloem replaced by Secondary Phloem
1. Structural differences difficult to study because of obliteration of earlier formed phloem
2. No clear growth rings in secondary phloem
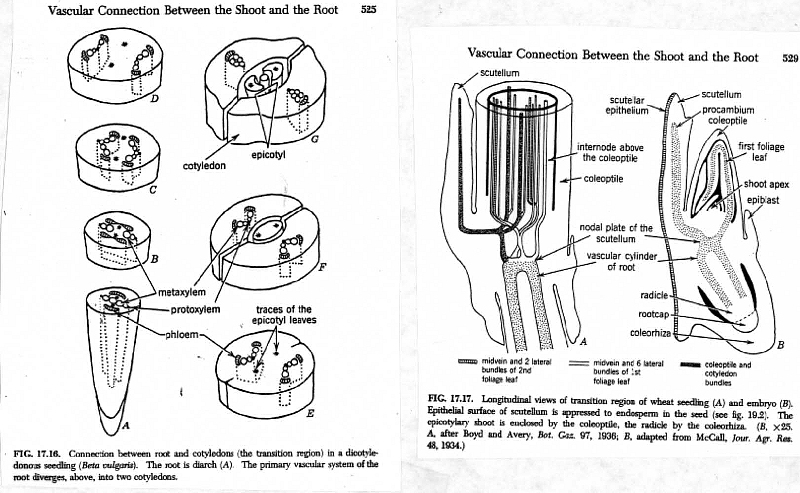
F. Overall pattern of organization of phloem parallels that of xylem in primary vasculature except in transition zone
II. Translocation of photosynthate
is from source to sink
| SOURCE | SINK |
| Storage Parenchyma of Endosperm & Cotyledons | Apical Meristems of Seedlings |
| Mature Photosynthesizing Leaves | Leaf or Floral Primordia in Shoot
Apex
Root Apical Meristems |
| Vascular Cambium | |
| Storage Parenchyma (Secondary Xylem, Fruits, Seeds) | |
| Storage Parenchyma (Secondary Xylem, Tubers, Corms, Bulbs) | Leaf Primordia in Shoot Apex
Root Apical Meristems |
| Vascular Cambium |
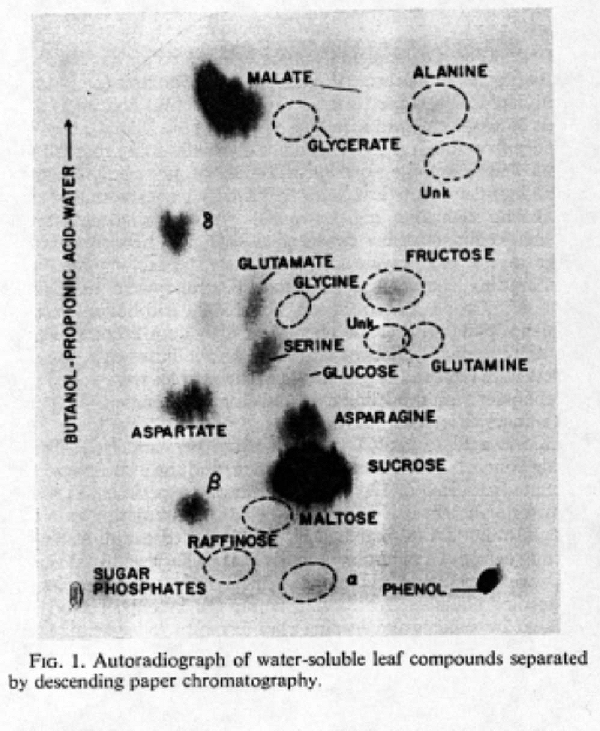
A. Typically studied using CO 2 constructed with radioactive carbon isotopes
B. Velocity of transport
1. 20 cm/h Pinus
2. 30 - 70 cm/h in dicot trees
D. Starch and or Cellulose converted to sugars before transportation
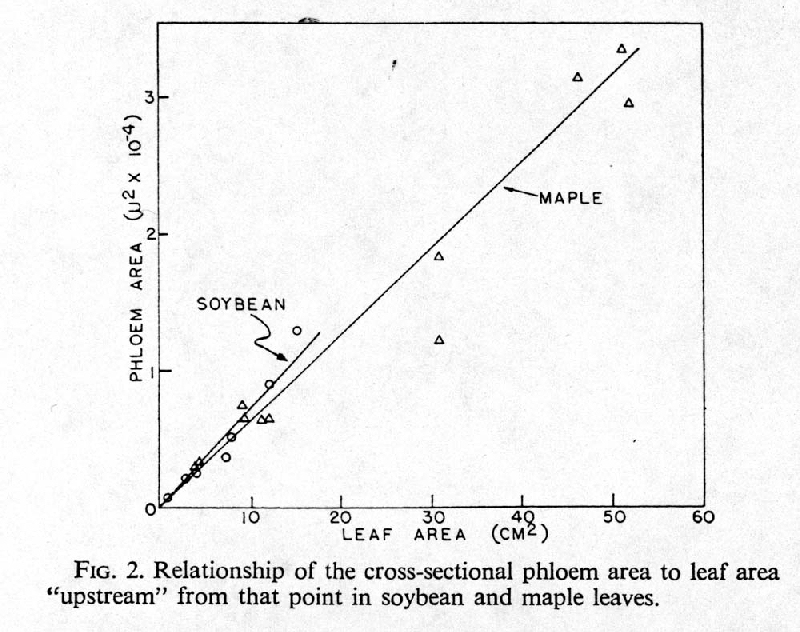
E. Appears to be a linear relationship between cross sectional area of phloem and upstream leaf area
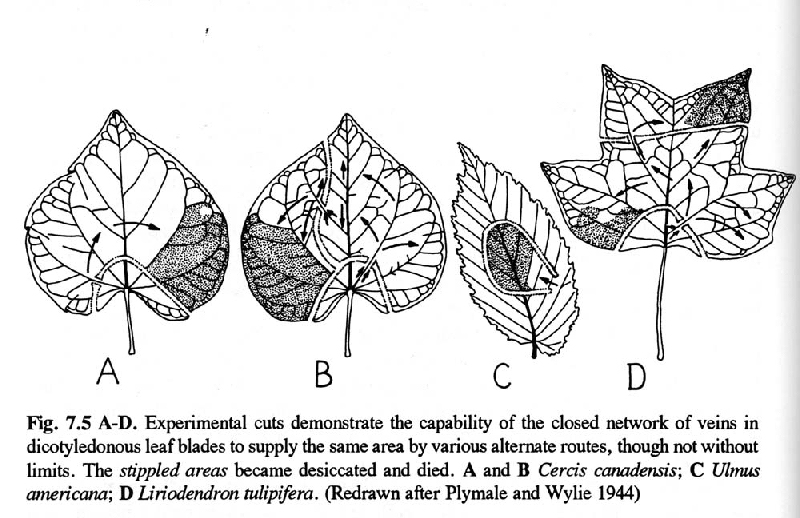
F. Leaf vein network provides alternative pathways for photosynthate translocation within constraints
II. Models of Phloem translocation
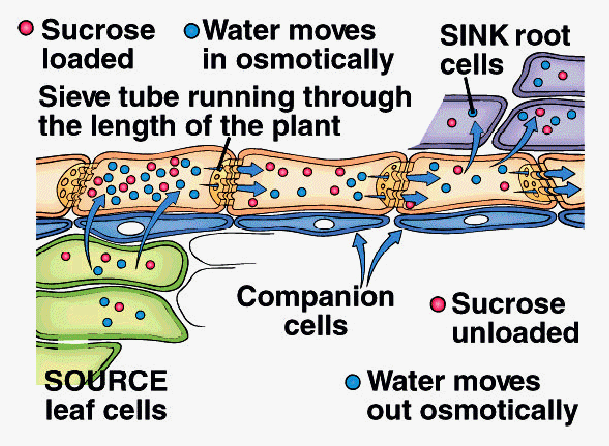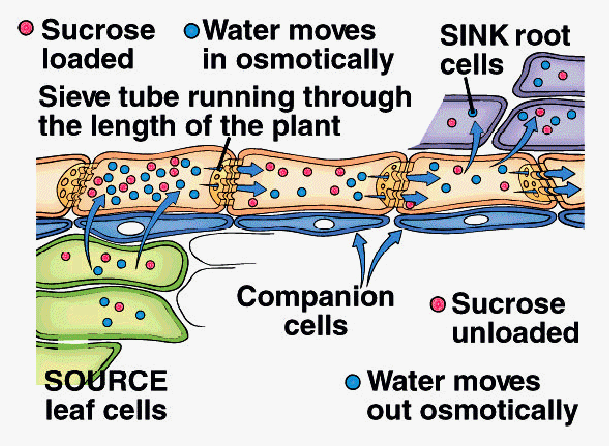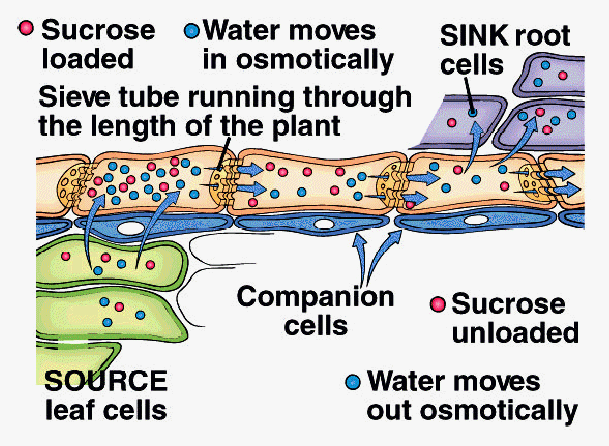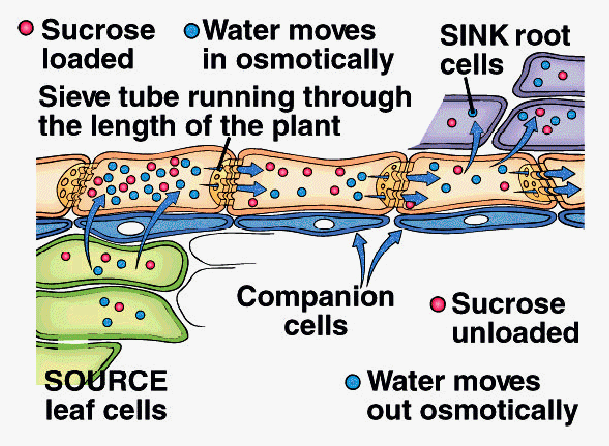
A. Pressure-Flow Hypothesis (E. Munch, 1927)
1. Sucrose actively loaded into sieve tube elements at SourceTissue
2. Water moves into STE by osmosis in response to increase in sucrose concentration
3. Hydrostatic pressure increases in STE as water moves in
4. Bulk flow of water and sucrose driven by hydrostatic pressure within ST toward Sink Tissue
5. Sucrose actively unloaded from STE into Sink Tissue
6. Water moves out of STE by osmosis into Sink Tissue
7. Translocation continues as long as pressure is maintained by active loading and unloading of sucrose and osmosis of water
B. Assumptions
1. Flow of water is laminar rather than turbulent
2. Laminar flow in capillary follows Poiseuille's Law
p = (8 * l * u * v) / ( p * r4) or v = (p * r4 * p) / (8 * l * u)
p = driving pressure required to move a fluid
with viscosity = u, at a
rate = v (ml per sec), through a
capillary of radius = r and
length = l.
p = (8 * l * u * v) / p * S ( n * r4)
S ( n * r4) describes attributes of a vascular bundle consisting of n sieve tubes
3. Laws of Conservation of Mass and Energy apply
4. Lateral membrane transport only occurs in source and sink regions
a. Even if they are open they introduce significant resistance into
the transport system
b. A pore 10 um in diameter would translocate100 times more photosynthate
than 1 um in diameter
C. Tests with
sugar beets
| PARAMETER | MODEL | EMPIRICAL |
| Velocity (cm/min) | 0.45 -1.48 | 0.9; 0.4 -1.9 |
| Concentration (%) | 8.3 - 28.0 | 8.8; 10 -25 |
| Osmotic Pressure (atm) | 6.0 - 20.1 | 18.04; 20 - 24 |
| Hydrostatic Pressure (atm) | 2.9 -16.5 | 15 - 20 |
| Specific Mass Transfer Rate (g/cm^2/h) | 7.3 | 4.8 ; 6- 18 |
a. Hydrostatic pressure, [], that can be generated by a given osmotic pressure, o, is less than rather than equal to the osmotic pressure, as had been assumed in the past
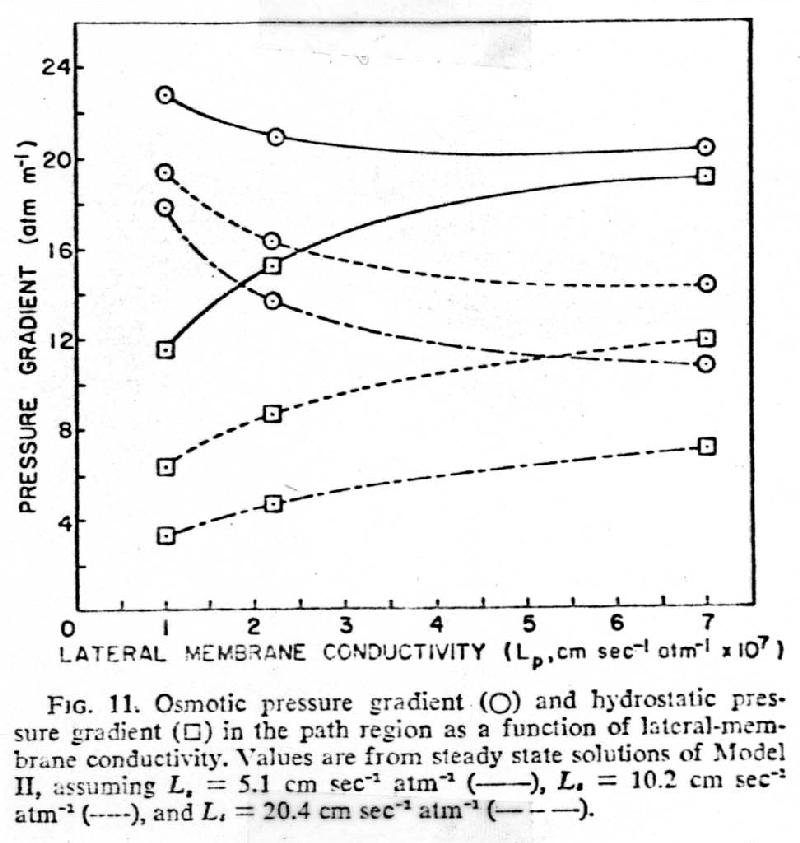
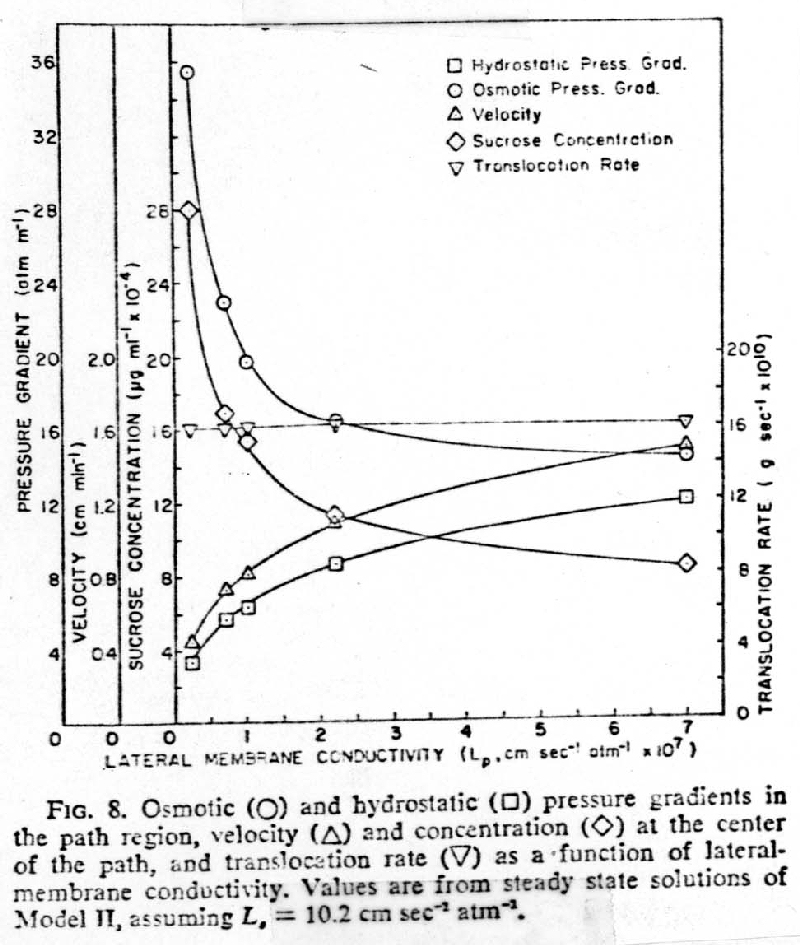
b. Lateral membrane conductivity is very important to the pressure-flow mechanism
1. At constant Lateral Membrane conductivity, Lp, an increase in conductivity of Sieve tube + Plates , (--- < - - - < -- - -- Ls), had little effect on the difference between hydrostatic and osmotic pressure gradients
2. With constant Sieve Tube + Plates conductivity an increase in Lateral Membrane Conductivity increases the hydrostatic pressure gradient, [], (because of increase in velocity,^) while decreasing both the osmotic pressure gradient , o, and the difference between the osmotic, o, and hydrostatic pressure gradient,[]
3. The osmotic pressure gradient must be sufficient to overcome the resistance of the lateral membranes, sieve tubes, and sieve plates
4. The hydrostatic pressure gradient must be sufficient to overcome
the resistance of the sieve tubes and sieve plates
|
|
|
D. Electro osmotic hypothesis
1. Requires a difference of electric potential across the sieve pore
a. Ionic pump in plasmalemma around sieve areas (costs more in energy than maintaining pressure gradient)
b. Traveling waves of conformational changes in microfilamentous
elements in pore is more efficient and would result in
traveling waves
of electrical change, which would facilitate flow through pores
2. Predicts sieve pores would be plugged
a.
Electro osmosis is faster through multiple small pores compared to few
larger pores because the most active zone in
each pore is the electrical
double layer at the interface between the solution and the pore wall